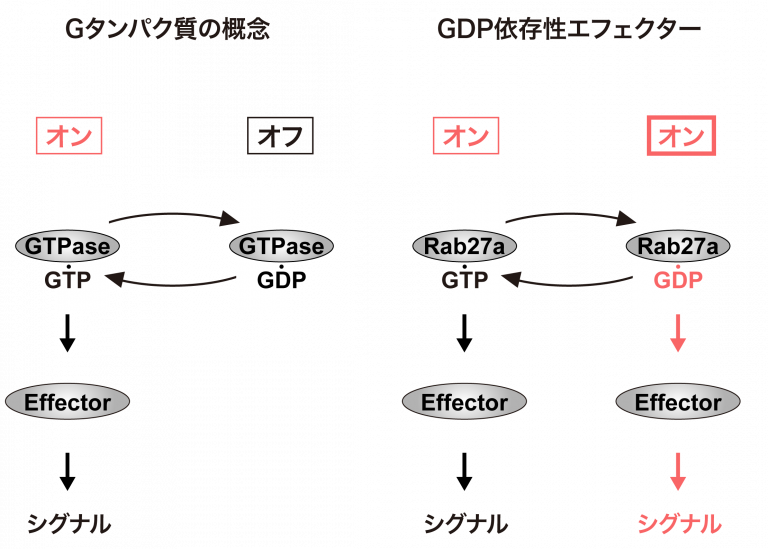
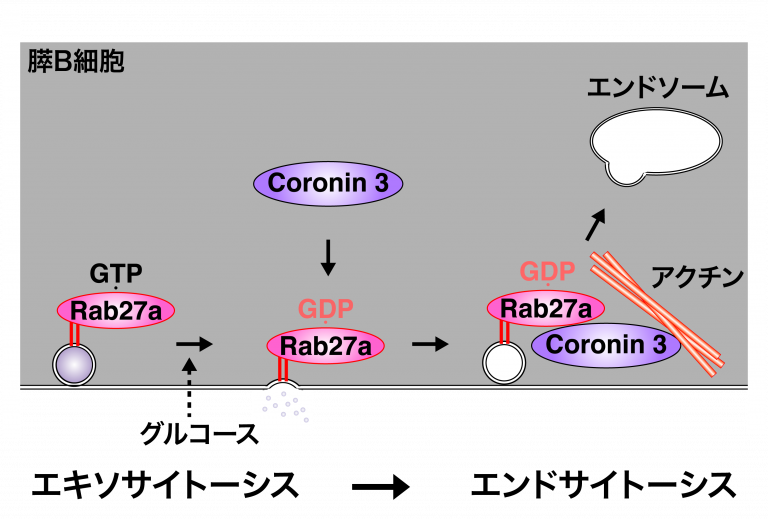
1. インスリン分泌制御機構の解析 近年の「科学技術重要施策アクションプラン」の重点的取り組みに掲げられているように、増え続ける糖尿病に対する新しい治療を開発することは、焦眉の課題です。我が国の糖尿病の大半を占める2型糖尿病の原因としては、膵B細胞からのインスリン分泌障害の占める割合が大きいことが臨床研究より明らかになっています。よって、2型糖尿病の病因を究明し糖尿病治療薬を創出するには、膵B細胞におけるインスリン分泌のメカニズムを明らかにすることが必要です。私たちは、分子スイッチであるGタンパク質に着目し、膵B細胞でインスリンを蓄えている小胞(インスリン顆粒)の動態が時間的・空間的にどのように制御されているのかを解析しています。 Gタンパク質は、GTP型が活性型(スイッチのオン)でGDP型が不活性型(スイッチのオフ)とこれまで考えられてきました。私たちは、GDP型のGタンパク質(Rab27a)に結合し、下流にシグナルを伝える分子(coronin3、IQGAP1)を世界に先駆けて発見しました。私たちは、「GDP型依存性エフェクター」の概念を提唱し、低分子量Gタンパク質ではどのくらい普遍的な現象であるのかを検討しています。 さらに私たちは、開口放出により細胞膜に融合したインスリン顆粒膜を再び細胞内に取り込むことで(エンドサイトーシス)、細胞の容積を分泌後も一定に保つメカニズムを明らかにしました。インスリン分泌刺激であるグルコースは、同時にエンドサイトーシスに関わるタンパク質群(coronin3とRab27a)を足場タンパク質(IQGAP1)を介して細胞膜近傍にリクルートします。リクルートされたcoronin3は、GDP型Rab27aと結合することで細胞骨格(アクチン)を束ね、エンドサイトーシスを制御することがわかりました。これは、GDP型依存性エフェクターのシグナル伝達を明らかにした最初の報告です。 グルコースによりRab27aがGDP型に変換される私たちの結果と合わせると、GTP型とGDP型間でのRab27aの「サイクリング」は、膵B細胞内での顆粒膜の「リサイクリング」と同期していることを示しています。インスリンの開口放出、あるいはその上流に介在する分泌機構に関する報告は枚挙にいとまがありません。しかし、エンドサイトーシスの制御はほとんど手がつけられていないと言って過言ではありません。私たちは、開口放出後のインスリン顆粒膜の動態とその制御機構の解明から、インスリンの新しい分泌制御機構を明らかにすることを目的としています。